



By Fred Reed
CONTROVERSIES
FRED REED • ORIGINALLY ON UNZ.COM, SEPTEMBER 8, 2016

Paleontologists have long thought that Archaeopteryx fossils, including this one discovered in Germany, placed the dinosaur at the base of the bird evolutionary tree. Recent evidence suggests the beast may have been a birdlike dinosaur. (Image credit: Humboldt Museum für Naturkunde Berlin)
|
This is atrociously long, criminally even, by internet standards but I post it anyway because I get occasional requests. Few will read it, which is understandable. Apologies. The Devil made me do it. Regular readers, if there is one, will have seen most of it before since in large part it is a gluing together of several columns.
“A scientist is part of what the Polish philosopher of science Ludwik Fleck called a “thought collective”: a group of people exchanging ideas in a mutually comprehensible idiom. The group, suggested Fleck, inevitably develops a mind of its own, as the individuals in it converge on a way of communicating, thinking and feeling. This makes scientific inquiry prone to the eternal rules of human social life: deference to the charismatic, herding towards majority opinion, punishment for deviance, and intense discomfort with admitting to error. Of course, such tendencies are precisely what the scientific method was invented to correct for, and over the long run, it does a good job of it. In the long run, however, we’re all dead, quite possibly sooner than we would be if we hadn’t been following a diet based on poor advice.” How the following Came AboutI was in high school when I began to think about evolution. I was then just discovering the sciences systematically and took them as what they offered themselves to be, a realm of reason and dispassionate regard for truth. There was a hard-edged clarity to them that I liked. You got real answers. Since evolution depended on such sciences as chemistry, I regarded it as also being a science. The question of the origin of life interested me. The evolutionary explanations that I encountered in textbooks of biology seemed weak, however. They ran to, “In primeval seas, evaporation concentrated dissolved compounds in a pore in a rock, a membrane formed, and life began its immense journey.” Still, I saw no reason to doubt this. If it hadn’t been true, scientists would not have said that it was. In those days I read Scientific American and New Scientist, the latter then still being thoughtfully written in good English. I noticed that not infrequently they offered differing speculations as to the origin of life. The belief in the instrumentality of chemical accident was constant, but the nature of the primeval soup changed to fit varying attempts at explanation. For a while, life was thought to have come about on clay in shallow water in seas of a particular composition, later in tidal pools with another chemical solution, then in the open ocean in another solution. This continues. Recently, geothermal vents have been offered as the home of the first life. Today (Feb 24, 2005) on the BBC website, I learn that life evolved below the oceanic floor. (“There is evidence that life evolved in the deep sediments,” co-author John Parkes, of Cardiff University, UK, told the BBC News website.”) The frequent shifting of ground bothered me. If we knew how life began, why did we have so many prospective mechanisms, none of which worked? Evolution began to look like a theory in search of a soup. Fifty-five years later in 2015, it still does. What Distinguishes Evolution from Other Sciences Early on, I noticed three things about evolution that differentiated it from other sciences (or, I could almost say, from science). First, plausibility was accepted as being equivalent to evidence. And of course the less you know, the greater the number of things that are plausible, because there are fewer facts to get in the way. Again and again evolutionists assumed that suggesting how something might have happened was equivalent to establishing how it had happened. Asking them for evidence usually aroused annoyance and sometimes, if persisted in, hostility. As an example, consider the view that life arose by chemical misadventure. By this they mean, I think, that they cannot imagine how else it might have come about. (Neither can I. Does one accept a poor explanation because unable to think of a good one?) This accidental-life theory, being somewhat plausible, is therefore accepted without the usual standards of science, such as reproducibility or rigorous demonstration of mathematical feasibility. Putting it otherwise, evolutionists are too attached to their ideas to be able to question them. Or to notice that others do question, and with reason. They defend furiously the evolution of life in earth’s seas as the most certain of certainties. Yet in the November, 2005 Scientific American, an article argues that life may have begun elsewhere, perhaps on Mars, and arrived here on meteorites. May have, perhaps, might. Somewhere, somewhere else, anywhere. Onward into the fog. Consequently, discussion often relies on vague and murky assertion, or ignores obvious questions. Starlings are said to have evolved to be the color of dirt so that hawks can’t see them to eat them. This is plausible and, I suspect, true. But guacamayos and cockatoos are gaudy enough to be seen from low-earth orbit. Is there a contradiction here? No, say evolutionists. Guacamayos are gaudy so they can find each other to mate. Always there is the pat explanation. But starlings seem to mate with great success, though invisible. If you have heard a guacamayo shriek, you can hardly doubt that another one could easily find it. Enthusiasts of evolution then told me that guacamayos were at the top of their food chain, and didn’t have predators. Or else that the predators were colorblind. On and on it goes. On any coral reef, a scuba diver can see, or rather not see, phenomenally good camouflage in creatures such as octopuses, said to prevent their being eaten. It does. But many fish are garishly colored. What is the advantage? Second, evolution seemed more a metaphysics or ideology than a science. The sciences, as I knew them, gave clear answers. Evolution involved intense faith in fuzzy principles. You demonstrated chemistry, but believed evolution. If you have ever debated a Marxist, or a serious liberal or conservative, or a feminist or Christian, you will have noticed that, although they can be exceedingly bright and well informed, they display a maddening evasiveness. You never get a straight answer if it is one they do not want to give. Crucial premises are not firmly established. Fundamental assertions do not tie to observable reality. Invariably the Marxist (or evolutionist) assumes that a detailed knowledge of economic conditions in the reign of Nicholas II substitutes for being able to answer simple questions, such as why Marxism has never worked. {This is a rather ridiculous objection by Fred, and, frankly, unworthy of him, but we'll et it pass, since his brilliance is demonstrated elsewhere and cumulatively.—Ed] This is the Fallacy of Irrelevant Knowledge. And of course almost anything can be made believable by considering only favorable evidence and interpreting hard. Third, evolutionists are obsessed by Christianity and Creationism, with which they imagine themselves to be in mortal combat. This is peculiar to them. Note that other sciences, such as astronomy and geology, even archaeology, are equally threatened by the notion that the world was created in 4004 BC. Astronomers pay not the slightest attention to Creationist ideas. Nobody does—except evolutionists. We are dealing with competing religions—overarching explanations of origin and destiny. Thus the fury of their response to skepticism. I found it pointless to tell them that I wasn’t a Creationist. They refused to believe it. If they had, they would have had to answer questions that they would rather avoid. Like any zealots, they cannot recognize their own zealotry. Thus their constant classification of skeptics as enemies (a word they often use)—of truth, of science, of Darwin, of progress. This tactical demonization is not unique to evolution. “Creationist” is to evolution what “racist” is to politics: A way of preventing discussion of what you do not want to discuss. Evolution is the political correctness of science. I have been on several lists on the internet that deal with matters such as evolution, have written on the subject, and have discussed evolution with various of its adherents. These men (almost all of them are) have frequently been very bright indeed, often Ivy League professors, some of them with names you would recognize. They are not amateurs of evolution, or high-school principals in Kansas eager to prove their modernity. I asked them questions, such as whether we really know what the primeval seas consisted of, etc. I knew the answers; I wanted to see how serious proponents of evolutionary biology would respond to awkward questions. It was like giving a bobcat a prostate exam. I got everything but answers. They told me I was a crank, implied over and over (again) that I was a Creationist, said that I was an enemy of science (someone who asks for evidence is an enemy of science). They said that I was trying to pull down modern biology (if you ask questions about an aspect of biology, you want to pull down biology). They told me I didn’t know anything (that’s why I was asking questions), and that I was a mere journalist (the validity of a question depends on its source rather than its content). But they didn’t answer the questions. They ducked and dodged and evaded. After thirty years in journalism, I know ducking and dodging when I see it. It was like cross-examining hostile witnesses. This is the behavior not of scientists, but of advocates, of True Believers. I used to think that science was about asking questions, not about defending things you didn’t really know. Religion, I thought, was the other way around. I guess I was wrong. The intent of this essay is not to debate with the ardent of evolutionism. To do so would be pointless. The problem is one of underlying set of mind, of why people believe and disbelieve things. The greatest intellectual divide is not between those who believe one thing and those who believe another, but between those who have an emotional need to believe something fervently and those who can say, “I don’t know.” The former group comprises those tedious Darwinists and Creationists who hurl imprecations at each other like fans of rival football teams. Each blockheadedly refuses to concede the slightest possibility that its doctrine might be other than infallible. To my mind they constitute the best evidence that we did not descend from monkeys, but have not yet ascended to them. Stupidity beyond a certain point is intractable. I write here for those who can look at the world with curiosity and calm, divining what can be divined and conceding what cannot, without regarding themselves as members of warring tribes. To judge by the writing on evolution in the public prints, there may be as many as three of these. “The universe is not only queerer than we suppose, it is queerer than we can suppose.” —J. B. S. Haldane “Queer”: Exactly the right word, suggesting more the world of Alice in Wonderland than the crisp, clean-edged, perfectly ordered and causal world of physics. This paradigm holds that existence is like a vast crossword puzzle. Some parts we have filled in, others we have not, but by its nature the puzzle is solvable, and it is only a matter of time before we know everything. This is awfully optimistic. Humans today are a puffed-up and overconfident species. We believe that we know everything, or shortly will. We have a sense of near-omniscience equaled only by that of teenagers. For do we not have smart phones and Mars landers and PET scans, and do we not all speak wisely of DNA? We are, if not gods, at least godlings on the way up. If you don’t believe this, just ask us. It was not always so. A thousand years ago, mankind cast a small shadow on the earth and lived in a dark and mysterious world. Little was known, about anything. Gods of countless sorts walked the earth. Spirits inhabited sacred groves. Lightning, the moon, the stars were…what? We had no idea. This brought humility. We now believe that nothing is or can be beyond our powers. A contemplative skeptic might advert to a few remaining details: We don’t know where we came from, why we are here, where “here” is, where we are going if anywhere, or what we ought to do. These are minor questions. We only think about them when we wake up at three a.m. and remember that we are not permanent. We are kidding ourselves. When people become accustomed to things that make no sense, they begin to seem to. Though we no longer notice it as we peck at tablet computers and listen to droning lowbrow shows about the conquest of nature, we still live in a weird and inexplicable universe, an apparently unending emptiness speckled with sparks of hydrogen fire. It is wicked mysterious. More things in heaven and earth, indeed. We are not as wise as we think. We are just smarter than anything else we know about. I reiterate Fred’s Principle: The smartest of a large number of hamsters is still a hamster. The Theory of Evolution is not just about biological evolution. It is part of a grand unified theory that seeks to explain everything (except things that it can’t explain, which it ignores). It runs briefly as follows: First came the Big Bang. Subatomic particles flew in all directions, coalesced into atoms and into molecules and stars. Planets formed, then oceans, and then life came about by chemical inadvertence. Evolution produced trilobites, dinosaurs, mammals, and us. In the popular version, though not in the scientific, evolution produces ongoing betterment. It is not particularly plausible. As someone said, evolution writ large is the belief that a large cloud of hydrogen will eventually turn into Manhattan. But, like a religion, it provides an overarching explanation of origins–the Big Bang–and destiny–we are getting better and better–and gives us a sense of understanding the world. In this it serves the purposes of a religion and is treated as such by its adherents. They react to questioning with anger and they see their hated opponents as Creationists–that is, adherents of another religion. Note that while in the Scopes Monkey Trial of 1925, Christian fundamentalists tried to outlaw Darwin, today evolutionists appeal to the courts to outlaw mention of Creation in the schools. This is not rational. Can anyone believe that describing Creation in high schools will deter students from studying biochemistry, and turn them into intellectual loin-cloth wearers burning textbooks? Interestingly, atheism has to be part of the evolutionist’s mental equipment since if any sort of god exists, or if there is life after death, or anything beyond the laws of physics, then these things might influence existence in a way outside of physics–and this cannot be allowed. Before going further, let us look at some of the questions ignored by evolutionism. In Evolution Writ Large nothing exists but physics. The Big Bang was physics, chemistry is the physics of the interactions of atoms, biochemistry a subset of chemistry and therefore also physics. Everything that happens in a cell is physics (to include biochemistry). Everything that happens in a living body, from movement to thought, is physics. Mutations are physical events. The behavior of DNA follows the laws of physics. Note that biological evolution is always regarded as an indivisible entity, yet in fact it consists of several distinct components that are logically separable. First, that life came about accidentally in the ancient seas (highly shaky and certainly not demonstrated). Second, that evolution occurred (as the fossil record would seem to show beyond reasonable doubt). Third, that natural selection drove evolution (demonstrable in some cases, plausible in a great many, and highly unlikely in yet others). Fourth, that random mutations drive natural selection (very shaky, but crucial to evolutionism). Fifth, that nothing else drives it. The unwillingness to recognize that these are separable leads to a tendency to believe that when one of them can be demonstrated–natural selection, say–it is regarded as confirmation of the whole edifice. It isn’t. An Embarrassing Necessity Before Getting to the Meat of Things Inevitably one who writes of evolution without being a PhD at CalTech is assaulted on grounds that he must be ignorant of practically everything. I claim to be an expert on nothing. However, I subscribe to the principle that most problems can be solved by the application of modest intelligence and obsessive-compulsive disorder. A fact forgotten today is that one can learn things by reading books. By doing so I have learned enough to talk about at least a few things, such as: Basophils, eosinophils, neutrophils. Descemet’s membrane, ciliary body, suspensory ligaments, retinal pigmented epithelium (the eye being of evolutionary interest). Peptide pituitary hormones, vasopressin and oxytocin. Osteoclast, osteoblast. Nephrons, glomerulus, Loop of Henle. Axon, dendrite, sodium in-potassium-out depolarization, neurotransmitters, receptor sites. Rough and smooth endoplasmic reticula, Golgi apparatus, lipid bilayers, hydrophobic and hydrophilic tails, lysosomes, ribosomes, epitopes, m-RNA, t-RNA, transcription, translation. Restriction enzymes, DNA polymerase. The Breeder’s Equation, selection differential, pleiotrophy, epistasis, narrow heritability.Purines adenine and guanine and pyrimidines cytocine and thymine (well, uracil in RNA). Degeneracy of the codon alphabet. Nucleotides, nucleosides, adenosine triphosphate, indels, mitochondrial cristae, single-nucleotide polymorphisms, polymerase chain reaction, restriction-fragment length polymorphism, electrophoresis. Luciferin, (and Luciferout?) luciferase, ATP. X chromosomal and mitochondrial DNA. Peptide bonds COOH to NH 2, water molecule extruded. Socially important compounds like 2, 4, 6- trinitrotoluene, toluene being benzene with a CH 3 group, bond resonance in benzene, pH, the negative log of the hydronium ion content. Levo- and dextro- isomers. Alkanes, alkenes, alkynes, al gore. Cambrian, Ordovician, Silurian, Devonian, Carboniferous, Permian. Purported transitional forms: The Ichthyostegids of, if memory serves, upper Permian sediments of eastern Greenland; Archaeopteryx, Bavaria 1861; coelacanth, Marjorie Latimer, sort of 1937 I think; and my favorite, Piltdown Man. The amniote egg. Saurischian and Ornothiscian dinosaurs. Sauropods, pseudopods, copepods. Etc. A matter that needs to be gotten out of the way before continuing is the insistence that, given billions of years–more accurately, about four billion–life had to from just because of all that time. This is by no means clear. In questions of the probability of complex events, time can mean surprisingly little. Consider the assertion famously made by James Jeans, often cited in connection with evolution, that a monkey typing randomly at a keyboard would eventually write all the books in the British Museum. This sounds plausible and, in a purely mathematical sense, is true. What are the odds? Consider a fair-sized book of 200,000 words that, by newspaper average, would contain about a million letters. To make it easy on the monkey, we will ignore upper case and punctuation and let him work with an alphabet of 26 letters. What are his prospects of getting the book in a given string of a million letters? The chance of getting the first letter correctly is 1/26 times the chance of getting the second letter, 1/26, and so on, making the chance of getting the entire book 1/261,000,000. Since 26 equals 10log 26, (log 26 being about 1.41) the chance of getting the entire book is 1 in 10log 26 x 1000,000 or about 101,400,000. Innocent looking numbers like this are remarkably intractable. For example, a billion billion monkeys (more monkeys than I want) typing a billion billion characters a second for a billion billion times the estimated age of the universe (1018 seconds ) would have essentially zero chance of getting the book. To give our monkey a fighting chance, let’s ask whether he would get even the title of a book, for example On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, which Microsoft Word tells me contains 119 characters. The monkey’s chance of getting the title in a given string of 119 is one in 10119 x x 1.41 or 10168 Thus our billion billion monkey at a billion billion characters a second for the life of the universe is essentially zero. Is the chance of accidentally forming a living Critter a similar problem? We don’t know, especially since evolutionists cannot tell us what the First Critter was. But it is their responsibility to tell us, first, what of what complexity formed and, second, why the odds are not astronomically against it. The point to take away is that the invocation of long periods of time can mean little when speaking of the probability of complex yet unspecified events. (1) Life was said to have begun by chemical inadvertence in the early seas. Did we, I wondered, really know of what those early seas consisted? Know, not suspect, hope, theorize, divine, speculate, or really, really wish. Bear in mind that chemical reactions depend crucially on molarity, pH, temperature, half-life of intermediates, and so on. The answer is, “no.” We have no dried residue, no remaining pools, and the science of planetogenesis isn’t nearly good enough to provide a quantitative analysis. 2) Do we know what conditions would be necessary for a cell to come about? No, we don’t. (3) Has the creation of a living cell been replicated in the laboratory? No, it has not. Here the evolutionist will say, “But, Fred, how can you repeat in the laboratory something that took millions and millions of years and billions and billions of gallons of sea water?” You can’t, but am I to believe it happened on the grounds that it can’t be proved? (4) Could it be shown to be mathematically probable that a cell would form, given any soup whatever? No, it couldn’t, and can’t. (At least not without cooking the assumptions.) (5) Have biochemists designed a replicating chemical entity that plausibly might have evolved into organisms such as we now have? No. 6) This next I ask, knowing that no answer is possible, to make a point: The more complex we postulate the First Critter to have been, the less likely that it would form accidentally. The less complex, the harder to explain why such a Critter has not been designed in the laboratory. With every passing year, the difficulty grows. In sum: If we don’t know what conditions existed, or what conditions would be necessary, and can’t reproduce the event in the laboratory, and can’t show it to be statistically probable, and can’t construct something that might have evolved—why are we so very sure that it happened? Would you hang a man on such evidence? To see the desperation of the search for plausible beginnings of life, look at this list, from the Wikipedia, of the wildly differing hypotheses, guesses, theories, and lunges, none of which have worked out. Does it give you a sense that evolutionists know what they are talking about? One hypothesis, as mentioned before, is that life swooped in from outer space on carbonaceous chondrites, or began on Mars (where it conspicuously has not been discovered by a platoon of itinerant Mars landers) and drifted to the earth. That is, life began where apparently there has never been life. The flexibility of evolutionary thinking is greatly to be admired. Here a point worth making briefly: The press often excitedly reports that “organic compounds” have been found on meteorites, or comets, or interstellar space, or in bottles of chemicals through which an electric spark has been passed. The unfortunate name “organic” suggests origins in living creatures, or the likelihood of turning shortly into living creatures. Actually, “organic chemistry” is, roughly, the chemistry of carbon chains. No living origins nor living intentions are implied. DDT is an organic compound, as is 2,4,6-trinitrotoluene, TNT. While evolutionists couldn’t demonstrate that life had begun by chemical accident, I can’t show that it did not. An inability to prove that something is statistically possible is not the same as proving that it is not statistically possible. Not being able to reproduce an event in the laboratory does not establish that it didn’t happen in nature. Etc. I didn’t know how life came about. I still don’t. Neither do evolutionists. Impossibility Theory and Common Sense, If Any If you look at evolution from other than the perspective of an ideological warrior who believes that he is saving the world from the claws of snake-handling primitive Christians in North Carolina, difficulties arise. Chief among these is the sheer complexity of things. Living organisms are just too complicated to have come about by accident. This, it seems to me, is apparent to, though not provable by, anyone with an open mind. Everywhere in the living world one sees intricacy wrapped in intricacy wrapped in intricacy. At some point the sane have to say, “This didn’t just happen. Something is going on that I don’t understand.” But an evolutionist cannot say that there is anything he can’t understand, only that there are things he doesn’t yet understand. Read a textbook of embryology. You start with a barely-visible zygote which, (we are told) guided by nothing but the laws of chemistry, unerringly reacts with ambient chemicals to build, over nine months, an incomprehensibly complex thing we call “a baby.” Cells migrate here, migrate there, modify themselves or are modified to form multitudinous organs, each of them phenomenally complex, all of this happening chemically and flawlessly on autopilot. We are accustomed to this, and so think it makes sense. The usual always seems reasonable. I don’t think it is. It simply isn’t possible, being a wild frontal assault on Murphy’s Law. Therefore babies do not exist. Quod erat demonstrandum. Unless Something Else is involved. I do not know what. Complexity upon complexity. In virtually invisible cells you find endoplasmic reticula, Golgi apparatus, ribosomes, nuclear and messenger and transfer RNA, lysosomes, countless enzymes, complex mechanisms for transcribing and translating DNA, itself a complex and still-mysterious repository of information. Somehow this is all packed into almost nowhere. That this just sort of, well, you know, happened is too much to believe. It began being believed when almost nothing was known about the complexity of cellular biology, after which, being by then a sacred text, it could not be questioned. And cannot. The foregoing is only the beginning of complexity. The many organs formed effortlessly in utero are as bafflingly elaborate as cells themselves. Consider (a simplified description of) the parts of the eye: The globe of three layers, sclera, choroid, and retina. Cornea of six layers, epithelium, Bowman’s membrane, substantia propria, Dua’s layer, Descemet’s membrane, endothelium. Retina of ten layers. Lens consisting of anterior and posterior capsule and contained proteinacious goop. The lens is held by delicate suspensory ligaments inside the ciliary body, a muscular doughnut that changes the shape of lens so as to focus. An iris of radial and circumferential fibers enervated competitively by the sympathetic and parasympathetic nervous systems in opposition. A pump to circulate the aqueous humor. On and on and on. And equally on and on for all the other organs, which last for seventy years, repairing themselves when damaged. 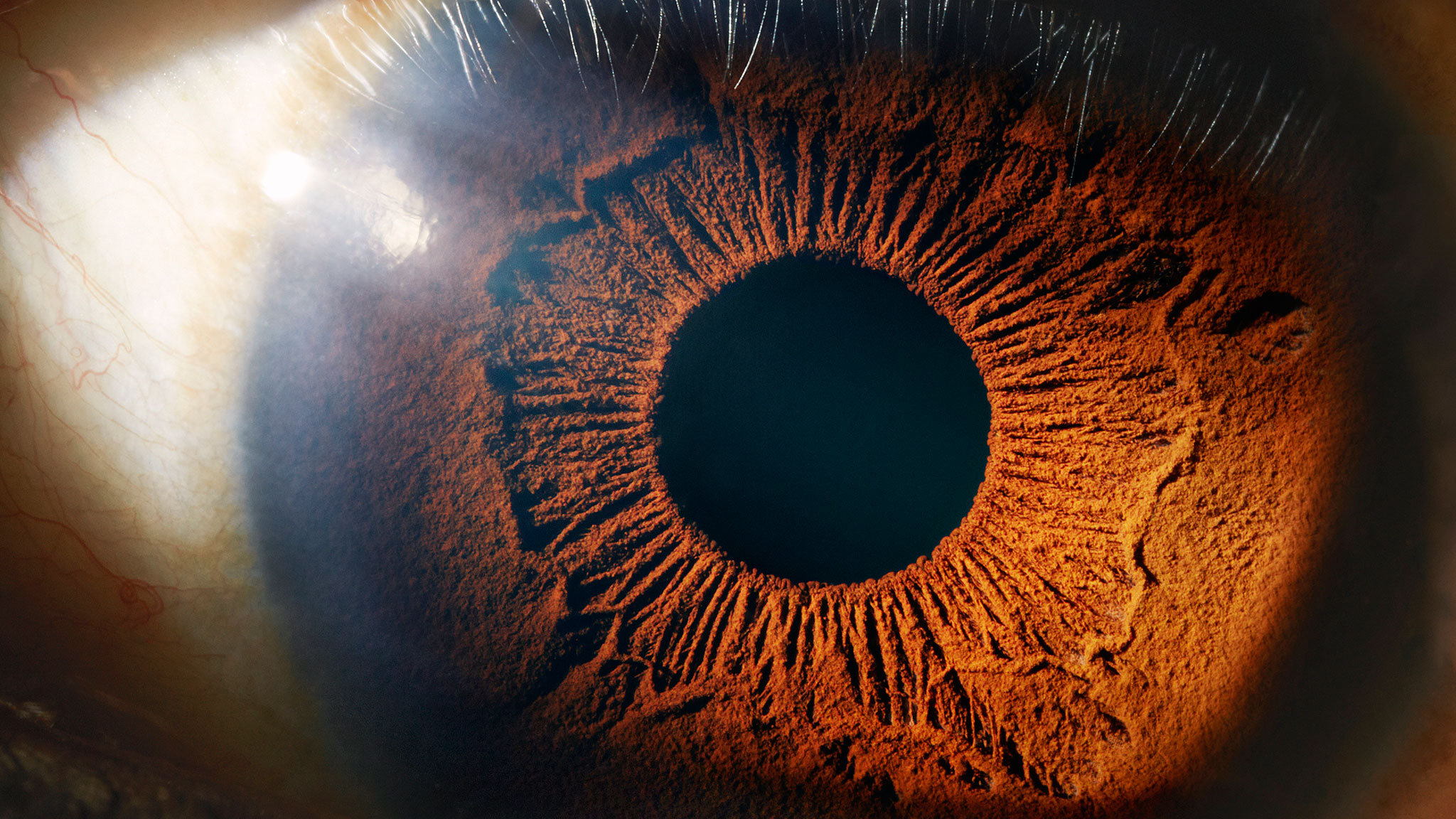 The human eye–or the eye, in general. Galactic complexity. Suspensory ligaments connecting the lens of the eye to the ciliary body. They form flawlessly on their own.I can’t prove that this didn’t come about accidentally. Neither can I believe it. The Details (Wherein Lurketh the Devil) At every level, complexity mounts. The following simplified description of the biochemical functioning of the retina is from Darwin’s Black Box: The Biochemical Challenge to Evolution by Michael Behe. The book, which I recommend, is accessible to the intelligent laymen, for whom it is written. The author includes the following technoglop to give a flavor of what is involved in vision. The sensible reader will skip through most of it. When light first strikes the retina a photon interacts with a molecule called 11-cis-retinal, which rearranges within picoseconds to trans-retinal. (A picosecond is about the time it takes light to travel the breadth of a single human hair.) The change in the shape of the retinal molecule forces a change in the shape of the protein, rhodopsin, to which the retinal is tightly bound. The protein’s metamorphosis alters its behavior. Now called metarhodopsin II, the protein sticks to another protein, called transducin. Before bumping into metarhodopsin II, transducin had tightly bound a small molecule called GDP. But when transducin interacts with metarhodopsin II, the GDP falls off, and a molecule called GTP binds to transducin. (GTP is closely related to, but critically different from, GDP.) GTP-transducin-metarhodopsin II now binds to a protein called phosphodiesterase, located in the inner membrane of the cell. When attached to metarhodopsin II and its entourage, the phosphodiesterase acquires the chemical ability to “cut” a molecule called cGMP (a chemical relative of both GDP and GTP). Initially there are a lot of cGMP molecules in the cell, but the phosphodiesterase lowers its concentration, just as a pulled plug lowers the water level in a bathtub. Another membrane protein that binds cGMP is called an ion channel. It acts as a gateway that regulates the number of sodium ions in the cell. Normally the ion channel allows sodium ions to flow into the cell, while a separate protein actively pumps them out again. The dual action of the ion channel and pump keeps the level of sodium ions in the cell within a narrow range. When the amount of cGMP is reduced because of cleavage by the phosphodiesterase, the ion channel closes, causing the cellular concentration of positively charged sodium ions to be reduced. This causes an imbalance of charge across the cell membrane that, finally, causes a current to be transmitted down the optic nerve to the brain. The result, when interpreted by the brain, is vision. If the reactions mentioned above were the only ones that operated in the cell, the supply of 11-cis-retinal, cGMP, and sodium ions would quickly be depleted. Something has to turn off the proteins that were turned on and restore the cell to its original state. Several mechanisms do this. First, in the dark the ion channel (in addition to sodium ions) also lets calcium ions into the cell. The calcium is pumped back out by a different protein so that a constant calcium concentration is maintained. When cGMP levels fall, shutting down the ion channel, calcium ion concentration decreases, too. The posphodiesterase enzyme, which destroys cGMP, slows down at lower calcium concentrations. Second, a protein called guanylate cyclase begins to resynthesize cGMP when calcium levels start to fall. Third, while all of this is going on, metarhodopsin II is chemically modified by an enzyme called rhodopsin kinase. The modified rhodopsin then binds to a protein known as arrestin, which prevents the rhodopsin from activating more transducin. So the cell contains mechanisms to limit the amplified signal started by a single photon. Trans-retinal eventually falls off of rhodopsin and must be reconverted to 11-cis-retinal and again bound by rhodopsin to get back to the starting point for another visual cycle. To accomplish this, trans-retinal is first chemically modified by an enzyme to trans-retinol— a form containing two more hydrogen atoms. A second enzyme then converts the molecule to 11-cis-retinol. Finally, a third enzyme removes the previously added hydrogen atoms to form 11-cis-retinal, a cycle is complete. I can perhaps imagine an Airbus 380 assembling itself. I cannot begin to imagine the foregoing evolving on its own. Or working flawlessly for more than a millisecond. Worse Than Intelligent Design: Layers of Impossibility If in an unexplored region of the Amazon Basin you find a grass hut next to a dugout canoe, you may not know who made them, but you suppose that someone must have. This is the theory of Intelligent Design. When you find in nature systems of unfathomable complexity that nonetheless work flawlessly, it is not unreasonable to suspect that they were designed, and perhaps sustained, by someone, or something. I have no idea who or what or why. Equally mysterious—equally impossible, I would say—is how biological systems can function at all, no matter how they came into being. The workings of every detail of, say, a human body can indeed be explained mechanistically, in terms of chemistry and physics, and this is the result that comes out of experimentation. In the laboratory you can show, or seem to show, that enzyme A binds to enzyme B, activating enzyme C and allowing enzyme D to do whatever enzyme D does. (You can show that a massive federal program makes sense in detail. But does it work in practice?) But to believe that 180 pounds of infinitely complex, interacting chemical reactions (me, for example) can go on for seventy years without utter collapse requires powers of belief beyond the wildest imaginings of religious faith. The whole is less possible than the sum of its parts. Something is going on that we do not understand. Consider a plane geometer. He deals with a limited domain of planes, lines, points, and angles, and nothing else. These produce elegant mathematics and useful results. He cannot deal with volumes, momentum, or tailgate parties, because these cannot be derived from the elements of his domain. They are beyond the scope of his subject. The domain of the sciences is physics, its elements being space, time, matter, and energy, however hyphenated. Everything in science ultimately reduces to physics. Evolution is the physics of interactions of biochemical systems with their physical environment over time, and thus also is a subset of physics. Nothing can happen in evolution that does not derive from and follow the laws of physics. Just as a baseball game cannot be derived from or be explained by plane geometry, which does not contain matter, energy, time, or space of three dimensions, neither can such things as thought, consciousness, morality, volition, or exaltation be explained by physics. The desire to strangle your mother-in-law does not fall out of the equations of motion. When evolutionists try to explain behavior such as altruism in terms of physics (which is what they are doing, though most of them don’t know it) they are like a plane geometer trying to explain a cheeseburger in terms of lines and angles in a plane. It can’t be done. The trouble with the sciences (though not with all scientists) is exactly this, that they try to explain within the domain of physics things that are outside of its purview. Studying Us: Explaining the Explainers The sciences get into particular difficulties when they try to explain the explainer, which is to say us. Consider the brain which, we are told, is just an electrochemical machine. Everything that happens in the brain, we are told, follows the laws of chemistry and physics. And this certainly seems to be the case. For example, neurotransmitters diffuse across the synaptic gap: pure chemistry and physics. They bind to receptors on the other side: pure chemistry and physics. Enzymes like acetylcholinesterase clear the residue from the gap: pure chemistry and physics. The resulting nervous impulse sails down the distal fiber as it depolarizes, sodium in, potassium out: pure chemistry and physics. It is as mechanical as a 1901 typewriter. Which means that the brain cannot, and thus we cannot, make choices. Physical systems cannot choose what to do. A bowling ball dropped from the top of the Washington Monument cannot decide to fall up, or sideways, instead of down, nor choose how fast to fall, nor how far. Similarly, the end point of a physical system is determined by starting conditions. A molecule of a neurotransmitter binds ineluctably to a receptor because of stereochemistry and charge. It cannot not bind. It follows then that we cannot choose one action over another. Our thoughts are predetermined by the physicochemical states of our brains. We think what we think because it is physically impossible to think anything else. Thus we cannot think at all. QED. Unless Something Else is going on. I don’t know what. Paradox is a consequence of domain bloat. Descartes famously said, “Cogito ergo sum.” Ambrose Bierce less famously but more insightfully said, “Cogito cogito, ergo cogito sum. Cogito.” Most people think that, “fitness” meaning “suitability for a purpose,” survival of the fittest means that the smarter, stronger, and faster survive and produce more offspring than the stupid, weak, and slow. It does not. The study of such things is called population genetics and, as a professor of it says, “In population genetics, fitness means the rate of successful reproduction, nothing else.” That is, fitness does not promote survival, but is survival. The circularity is well known: Why do they survive? Because they are fit. How do you know that they are fit? Because they survive. If fitness means the rate of successful reproduction, we encounter the interesting conclusion that a woman with a genetic IQ of sixty and twelve retarded children by forty-five drive-by fathers is more fit than a Harvard math professor who runs Triathlons but has two children. If instead of “fitness” with its almost inescapable overtones of “superiority,” we used “reproduction rate,” clarity would follow. Perish forbid. A staple of evolutionism is that evolution works to maximize the number of offspring, thus passing on successful genes. This is plausible but, in the case of us, counter to observation (but why let facts debilitate a perfectly good theory?) The populations of advanced countries, all of which could easily support larger numbers of people, are actually falling. For example, Japan, Spain, Italy, Germany, and Russia. In Mexico, as the standard of living rises, the birth rate falls sharply. How one passes on one’s genes by not passing them on is a mystery of population genetics. Meanwhile the populations of black Africa, the civilizational equivalents of the unwed mother with an IQ of 60, grow rapidly. Which is to say that in advanced countries, reproduction of individuals is inversely proportional to circumstances favoring it–intelligence health, wealth, and education. Among nations, as noted, a similar phenomenon exists. When this is pointed out, evolutionists hem and haw (or should I say hem and her?), sometimes say that evolution no longer applies to humans, (though they simultaneously insist that evolution is ongoing and rapid) and then often blame falling populations on contraception, as if this were an outside force, like drought or a new predator. But saying that contraception causes falling populations ls like saying that spears cause hunting. People wanted to eat, so they invented spears. They wanted not to have children, so they invented contraception. Not passing on one’s genes is now almost a preoccupation. Another peculiarity is populational altruism. Countries with declining populations intentionally import inferior but more-fecund genetic groups. Sweden for example imports black Africans. In the United States, the white population feeds and clothes huge number of genetically utterly distinct blacks, and actually seems to be growing them. The Darwinian advantage of this is elusive. Current Human EvolutionEvolutionists insist that human evolution continues today at a rapid pace. There is nothing illogical in this to the extent that it is a matter of selective breeding and that evolution is defined as a change in phenotype. In some cases it can be shown to happen. Consider for example cognitive stratification, in which very smart people tend to go to Ivy universities, marry each other, and produce smart children. The children will tend to revert toward the mean but, as they interbreed, the mean will rise. Thus a fairly distinct subpopulation comes about. While such things certainly can occur, problems arise in the evolutionists’ casual attribution of traits to evolutionary change. The first is that “selective pressure” usually cannot be measured and cannot be correlated with its purported results. Traits are regularly attributed to genes that have not been demonstrated acted upon by selective pressures that cannot be quantified to produce results that cannot be correlated with the pressures. The second is that results often seem to be inversely related to what would seem to be obvious selective advantage. Often it seems that evolution is driven less by selective pressure than by the absence of selective pressure. Before the advent of modern medicine, people with inferior genetic endowments– low resistance to disease, or possession of genetic diseases such as diabetes, serious retardation, etc.–tended to die before reproducing. This selective pressure served to keep those diseases at a low level in the population. Today the defective are kept alive to reproductive age, have children, and thus rapidly increase the prevalence of those diseases in the population. There is the curious fact that traits of very little obvious value flourish, while those seemingly important do not. Consider the epicanthic fold, which makes the Japanese and Chinese slant-eyed. Evolutionists I have read assert alternately that the fold serves to conserve energy or to protect the eye against icy winds, thus furthering survival. Characteristically, they cite no studies demonstrating that the fold does either of these things: In evolution, plausibility substitutes for evidence. The fold has become universal in the populations, suggesting that powerful selective pressures must have been responsible. But what pressures? Do we really believe that the fold provides enough protection to the eye, if it provides any at all, to result in its possessor having more children than others? Do foldless Vikings go blind? Where is the evolutionary noise level? At what point is the selective advantage, if any, so slight as to make no difference? Which brings us to a baffling question. Why does a trait with very little or no reproductive value–the fold–become universal, when traits such as high intelligence, great physical prowess, astonishing eyesight, and so on did not become even common? The genes for all of these already exist in the population without the need for mutations. If traits that conduce to reproduction become ever more prevalent, it follows that traits that do not become prevalent do not conduce to reproduction. These would seem to include the aforementioned–intelligence, strength, and so on–as these seem no more common now than in classical antiquity. If human evolution continues today at a rapid pace as evolutionists say (and indeed it may) it follows that selective pressures must be fairly intense. It is reasonable to ask, what pressures to what end? Cognitive stratification–the self-selection of people with IQs of perhaps 130 and up–qualifies and may lead to a blurry-edged yet distinct subpopulation. Yet pressures would otherwise seem to be low now. In modern human populations, in which almost no one dies in infancy, almost everyone marries, and almost everyone has the same small number of children, the number of offspring is not determined by life-or-death selection. The football captain gets the prom queen, but the class nerd gets the nerdette and can have as many children. Almost everyone lives past reproductive age, so there is little culling effect as the slow are eaten by wolves. The genetically sickly are kept alive and allowed to reproduce by medicine. Consequently it is hard to imagine Darwinian selection occurring with much ferocity. Nor can I see evidence for more than minor changes in the 2500 years since Fifth-Century Athens. Statues by Phidias and Praxiteles and later Roman copies show people exactly like us. It is impossible to give IQ tests to the long dead, but Plato and Archimedes seem very like the best minds of today, and the writing of such as Xenophon are indistinguishable in complexity, clarity, and quality of mind from good modern writers. Nothing suggests that the ancients were any less athletic, bellicose, or agile than we are, or that they had senses any less acute. The 2500 years of rapid evolution appear to have produce a net of zero. The Bugs in DarwinA Thing is Not Possible Merely Because It Happens: The Tarantula Hawk It is easy to imagine how a complex system, once in existence, can, within limits, evolve under the influence of selective pressures. Any dog breeder can demonstrate this. Or think of the path from Eohippus to Clydesdale. The difficulty lies in knowing how the system came about in the first place. Consider the Tarantula Hawk, a gigantic wasp that begins life as an egg inside a paralyzed and buried tarantula, where its mother put it. This may seem unmotherly, but there is no accounting for taste. The egg hatches. The larva feeds on the spider, somehow knowing how to avoid the vital organs so as to keep the monster alive and fresh. It pupates and then, a new adult, digs its way out of the burrow.  Tarantula hawk feeding in hand. The sting is paralyzingly painful. Off it flies. Never having seen another wasp, or anything else, it finds one, and knows how to mate. (Mating, if you think about it, is a rather more complex process than it may seem to high-schoolers. Some insects mate while flying, which compounds the trickiness. Think airline pilots and stewardesses.) Never having seen a tarantula, it knows how to find one, knows that it needs to attack it, knows exactly how to sting it, knows that it must drag it to its burrow, which it knows it has to dig, knows how to lay its egg on the tarantula and how to bury it. Now, some of this may be imagined as evolving by gradual steps (emphasis on “imagined,” which in matters evolutionary is good enough) as required by Darwin. All it takes is enough time. In enough time, anything desired will happen. Of millions and billions of eggs deposited in unfortunate tarantulas, over millions of years, some larvae ate the spider’s vital organs and so died in a rotting spider, not passing on their genes. Others pupated but tried to dig out by going downwards or sideways, thus dying and not passing on their genes. Only those with don’t-eat-the-important-parts mutations and this-way-is-up mutations survived, and so their genes became universal. This we are told. But…but knowing what a tarantula looks like when you have never seen one, or seen anything, knowing that you need to sting it and just how, that you need to dig a burrow and drag the spider to it, and cover it up, when all of this has to occur in order or the whole process fails…. The Bot fly is a squat, ugly, hairy fly that (in one version anyway) catches a mosquito, lays its eggs on on said mosquito after positioning it correctly, and attaches them with a kind of glue. It releases the mosquito. When the little feathery syringe lands on, say, a human, the eggs drop off, hatch, and burrow into the host. These make nasty raised lumps with something wiggling inside them. Later the larvae exit, fall to the ground, and pupate. How did this evolve? Did a grab-a-mosquito gene occur as a random mutation (assuming that a single mutation could cause such complex behavior)? It would have to be a grab-a-mosquito-but-don’t-cripple-it gene. That is an awful lot of precise behavior for one mutation. At this point the bot fly would have a mosquito but no idea what to do with it. It would need simultaneously to have a stick-eggs-on-mosquito mutation. This would seem to require another rather ambitious gene. Catching the mosquito without laying the eggs, or squashing the mosquito in the process, or laying eggs in mid air without having caught the mosquito, would seem a losing proposition. None of these awfully-lucky mutations would be of use without the others. How do you evolve this elaborate dance by gradual steps?  Close up of giant hornet Vespa mandarinia japonica Living things are impossible, but some are more so. Consider brains. Larger brains supposedly allow more-complex behavior. In a laptop civilization, we refer to this as “processing power.” But consider hornets, cautiously. These have very complex behavior but almost no brains or other nervous tissue. Yet their unbrains control six multiply-joined legs (any robotics engineer will tell you that this is a massive problem), and allow them to fly precisely, also a very difficult problem. They know just how to chew wood fiber to make a paste from which they know how to construct complex nests. They know how and when to mate, which is not a simple a process. The same barely existent nervous system operates various senses and interprets the resulting data, which also isn’t easy. They find food, inform the others of its location, and navigate effortlessly over long distance.  Ant Camponotus ligniperdus Even worse than hornets: Here you see a very little ant, and very little of very little ant consists of nervous tissue. Yet they too build nests, control legs, and senses, and digestive organs, find food, care for young, and lots more. It takes an evolutionist not to suspect that something is going on that we do not understand. Another ant might notice, but not an evolutionist. Yet hornets are pointy-headed intellectuals compared to pharaoh ants, above, those super-tiny picnic abominations of which several would fit on a hornet’s eye. They too have complex social organization and so on—with hardly any neurons. In general, the behavior of social insects is probably more complex than that of whales. It is inexplicable, or at least unexplained. Metamorphosis: You Can’t Get There from Here Straight-line evolution, for example in which Eohippus gradually gets larger until it reaches Clydesdale, is plausible because each intervening step is a viable animal. Darwin himself pointed this out. In fact it is just selective breeding. Yet many evolutionary transformations seem to require intermediate stages that could not survive. Metamorphosis in insects is perhaps the most baffling example. Consider. There are two-cycle bugs that lay eggs that hatch into tiny replicas of the adults, which grow, lay eggs, and repeat the cycle. The four-cycle bugs go through egg, larva, pupa, adult. Question: What are the viable steps needed to evolve from two-cycle to four-cycle? Or from anything to four-cycle? Here I am stumped. As best I can see, the eggs of the two-cycler would have to evolve toward being caterpillars, which are enormously different structurally and otherwise from adults. Goodbye legs, chitinous exoskeleton; head, thorax, and abdomen, on and on. Whatever the first mutation toward this end, the resulting newly-hatched mutant would have to be viable—able to live and reproduce until the next mutation occurred. Let us consider this question carefully. We begin with a two-cycle bug, that for convenience we will call a roach, which will endeavor to evolve into a bug that, also for convenience, we will assume to be a butterfly. The roach has the insect’s standard body plan of head, thorax, and abdomen, and the usual chitinous exoskeleton. From a spirit of charity we will assume that it is a flying roach to give it a head start toward butterflyhood. To achieve that exalted end, our roach would first have to evolve into a caterpillar–that is, a larval form. It is difficult to see how this could occur at all, or why. To become a caterpillar, our roach would have to lose its jointed legs, exoskeleton, and body plan. Since not even the most hopeful evolutionist could attribute such sweeping changes to one mutation, the transformation would have to proceed by steps involving at least several and probably many mutations. Losing the exoskeleton would leave it unarmored and unable to walk, not an obvious selective advantage. Or do we believe that head, thorax, and abdomen first merged mediated by a long chain of accidental mutations under mysterious selective pressures , and then it lost its exoskeleton and became, well– bait? But if these things did happen, they would lead to a free-standing race of caterpillars, a new species, necessarily being able to reproduce. Then, for reasons mysterious to me, these would have to decide to pupate and become butterflies. And the butterfly would have to lay eggs that became caterpillars. Which could not possibly work. Metamorphosis from caterpillar to butterfly is enormously complex and if you don’t get it right the first time, it’s curtains. It would depend on a great many steps which would have to appear simultaneously. First, our caterpillar would have to use its spinnerets (of mysterious provenance, but never mind) to make a cocoon, in which it would proceed to die because it hadn’t yet evolved metamorphosis. Why a caterpillar would think of doing this is not clear. To turn successfully into a butterfly, it would need the biochemical machinery to transform a mushy, legless, wingless, head-thorax-abdomenless worm into an utterly different creature. Where would it have gotten the impossibly complex genetic blueprint of the butterfly? Methinks something is going on that we do not understand. Note that the questions posed by these bugs are not merely pleasant musings on a slow afternoon. Either the Theory of Evolution can explain them, or the theory fails. The problem is usually referred to as that of Irreducible Complexity, the requirement that a great many mutations each of no value in itself, or actually harmful, appear simultaneously to create a given outcome. This term, implied in the foregoing, refers to the frequently observed existence in living organisms of systems that depend for their functioning on the simultaneous presence of things that would be either useless or detrimental by themselves, and thus make no evolutionary sense. For example, none of the individual steps of the bot fly’s complicated behavior with its mosquito would be of any value unless all the others were also present. This is irreducible complexity: take away any part and the system fails. Evolutionists insist that irreducible complexity does not exist. If this is true, then any biological system can be simplified step by step back to its origins without producing intermediate stages that could not survive. In particular, a living cell, the functionally important parts of which seem irreducibly complex, can be simplified bit by bit to produce the original First Critter. Why has this not been done? I find it interesting to imagine just how the simplification might be carried out. Perhaps by reducing the number of nucleotides per codon from three to two? This would allow coding only sixteen amino acids with no STOPs or STARTs. Can we eliminate transcription and go direct to translation? Get rid of the cell membrane so that everything inside floats off in different directions without extinguishing what was left? Surely this process would solve the problem of seeing how the cell evolved from the First Critter, and what that critter was. If there is no such thing as irreducible complexity. Traits often arise for which there is no good evolutionary explanation. Evolutionists here have two escape hatches, (1) conservation of energy, and (2) sexual selection. For example, if one points out that humans are weak and would be more survivable if they were as strong as, say, chimpanzees, the response is that having larger muscles would require a higher caloric intake to maintain them, and lead to starvation if there were a drought. Sexual selection: If peacocks have hugely conspicuous tails that would attract predators, the explanation is that all the girls love a good tail, so the guy leaves more children. Let’s look at these notions. Conservation of energy. Human beings are conspicuous in the natural world for being weak and slow, and for having poor senses of smell and hearing. Why? Evolutionists have multiple stories. One is that because humans walk upright, they can see farther on open veldt and thus have substituted vision for other senses that just are not necessary. This makes no sense which, as so often in matters evolutionary, doesn’t matter. Obviously being able to detect approaching predators at night by smell would be a great advantage. Lions are the color of dirt and dead vegetation and take advantage of both. Horses, which have good vision, and eyes at about the level of a human’s, have an excellent sense of smell. This story doesn’t live up even to the usual evolutionary standard of vague plausibility. Another explanation of the poor olfaction of humans is that a more acute sense would require larger olfactory regions in the brain and, since a surprisingly large proportion of the body’s energy is expended by the brain, these larger olfactory regions would increase the need for food and cause starvation in time of famine. Consider. Rats have a much better sense of smell than do humans, which they use in finding what they regard as food. A rat’s brain weighs two grams, a human’s about 1350. Let us assume that a rat's entire brain is dedicated to smell, which of course it isn’t. Adding all of a rat’s brain to the human would increase its size from 1350 to 1352 grams, an increase of 2/1350 or .15%, Since the brain uses 15% of a human’s energy budget, the overall increase in energy requirements is 2/1350 X 100 X .15, or .02%. Not 2%, but .02%. This minute increase cannot possibly offset the advantages of an acute sense of smell. The same reasoning applies to other senses, such as hearing. And of course people already have olfactory regions. They just don’t do much. Sexual selection Another way of explaining things that otherwise make no sense is “sexual selection.” Many things would seem to work against survival, yet persist in nature: huge antlers not usable in combat, the gorgeous tails of peacocks, and large breasts in humans, among many others. Why do women have conspicuous breasts? They are not needed to produce adequate milk, and they are a substantial physical disadvantage in running (thus we have sports bras). One would expect them to disappear.  Darwin These things are useless for defense as the animal would have to stand on its head to present them to an enemy. They cannot help balance or speed. We are to believe that they serve as sexual attractants because otherwise they are inexplicable. (I favor option B.) Since it is unlikely that headgear so glorious sprang from a point mutation, they must have begun as mere bumps. And all the girls swooned? The answer is sexual selection: men are attracted to large breasts, so those women with them mate and have more children. This suggests that women with modest endowments will have trouble getting laid, which in turn suggests that evolutionists need to get out more. The problems with sexual selection are twofold. First is that sexual selection requires a pre-existing attraction to large breasts. Otherwise in a cave society when the first woman through mutation appeared with big ones, we would hear one cave man say to another, “Geez, Urk Urk, what’s wrong with Sally?” “Beats, me, Ralph. Maybe it’s cancer.” But why would there be a preference for large breasts when there were no large breasts to prefer? The second problem is that if sexual selection favored large breasts, by now most women would have them, which visibly is not the case. (Again, compare Greek statues of 2500 years ago look like us.) And of course when the sexually-selected trait became general in the population, it would cease to be of advantage. While consciousness seems the defining characteristic of life, (“I am conscious, therefore I am.”) or at least of the higher forms of animal life, it cannot be derived from physics. It cannot even be detected. Are ants conscious–or, for that matter, rocks? Are dogs less conscious than people, and ants less conscious than dogs.? Or are they just less intelligent? How could we tell? The questions may seem silly, but they are not. They are tied up with our ability to make decisions, which physics says we cannot. Again, our brains, which are physical systems, cannot act on decisions any more than a dropped bowling ball can decide to fall sideways. Here is something outside of physics, and therefore outside of evolution, which must be ignored, and is. When people have engaged in bitter ideological war over a theoretical ship dear to them, they tend to overlook the cracks and stains and leaks in the planking. Evolutionism is full of such. An unaffiliated skeptic can point them out in droves. In evolution, traits which conduce to survival, and thus to the passing on of genes, are supposed to flourish, while traits that work against this happy passing on, or simply do nothing, are supposed to be eliminated. Does this happen? Often, yes. Not infrequently, no. An obvious problem is male homosexuality. Homosexuals seldom have children. How does not passing on one’s genes contribute to passing on one’s genes? The condition would seem to be a prime candidate for elimination by evolution, yet it has apparently been with us forever. If this cannot be explained away, then something is wrong with the theory in at least this case. Here evolutionists fall back on their Maginot Line, vague plausibility. For example Greg Cochran, a physicist of immoderate pomposity at the University of Utah, says that a virus causes homosexuality. The evidence for this virus? Homosexuality. Yet the chief characteristic of the virus unfortunately seems to be undetectability: No one can find it. Without this virus, the evolution would fail, at least at this point. Therefore a virus must, must, must exist. We infer reality from the needs of our theory. Other reproductive traits suffer from similar inexplicability: what are the reproductive value of suicide, masochism, sadism, schizophrenia, and so on? Should these not be filtered from the gene pool? Must we invoke viruses to explain these too? Schizophrenia: A Neanderthal who thought that the CIA put transmitters in his teeth and tried to shake hands with Kodiak bears might limit his reproductive opportunities. While a suicide who blows himself up with a bomb may be said to be disseminating his DNA, it serves little reproductive purpose. Yet all of these things have been with us forever. I therefore propose the existence of a virus for each of these peculiarities. And perhaps one for sun spots. Again, the problem is Domain Bloat, insisting that one’s theory explains what it can explain but also what it can’t. Next, consider pain. If you step on broken glass, it hurts, so you stop doing it, and don’t end up crippled and eaten by wolves, and so you can pass on your genes upon encountering an amiable maiden. This makes sense. What doesn’t make sense is the agonizing pain caused by many circumstances about which, pre-medicine, the victim could do nothing. Kidney stones, for example, are paralyzingly painful. A choroidal hemorrhage, behind the retina, is hideous. The agony has no utility since the premodern sufferer could do nothing about it. For that matter, the contribution of migraines to survival is not apparent, as a person rolling on the ground and clutching his head would seem vulnerable to ingestion. On and on. Why the abundant pain receptors with no function? Why do they not, like Marx’s state, wither away? Perhaps instead of asking, “How does evolution explain a thing?” we should occasionally ask “Does evolution explain it?” Impossible, Impossibler, Impossiblest Clear examples of things outside the domain of physics are morality, right and wrong, Good and Evil. A Darwinist cannot say that some things are intrinsically wrong. “Wrong” cannot be derived from physics. Instead he must show that moral behavior exists because it promotes the passing on of genes. Thus I nurse my brother back to health when he has a broken leg because together we can protect ourselves and our women better and thus pass on our genes. This of course runs into all sorts of problems. In Moslem countries, “honor kills” are thought acceptable: killing one’s daughter on discovering that she had engaged in sex before marriage (thus offering to pass on her, and her father’s, genes, but never mind). In Christian countries, this is called “first-degree murder,” and likely results in Dad’s sitting in a funny chair with wires running to it. Are we to believe that Moslem genomes contain a kill-daughter gene? Or is the obvious explanation, culture, to blame? It is interesting that evolutionists do not believe their own doctrine. Suppose a Darwinist found out that my hobby was using a blowtorch to torture to death children with severe genetic retardation. He would be horrified. “Why?” I would ask. “We certainly do not want them passing on their extremely defective genes. Caring for them expends resources that would be better spent in raising more children to pass on our genes. Torturing them has no more evolutionary meaning than killing them instantly. Actually, all I am doing is terminating certain chemical reactions and allowing others to begin. What then is your objection?” His objection would of course be that torturing children is wrong. But, again, “wrong” doesn’t exist within the domain of physics, and so of Darwinism. Domain bloat.  Fred A. Reed is a journalist, author, and translator who has published and translated several books. He is a three-time winner of the Governor General's Award for French to English translation, for his translations of works by Thierry Hentsch and Martine Desjardins, and received six further shortlisted nominations. Fred is a happy expat residing in Mexico with his Mexican wife. Fred A. Reed is a journalist, author, and translator who has published and translated several books. He is a three-time winner of the Governor General's Award for French to English translation, for his translations of works by Thierry Hentsch and Martine Desjardins, and received six further shortlisted nominations. Fred is a happy expat residing in Mexico with his Mexican wife. |
[premium_newsticker id="211406"]
AUTH

This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
The views expressed are solely those of the author and may or may not reflect those of The Greanville Post
All captions, quotes, annotations and pullquotes by the editors not the authors.
![]()